 |
Главная |


Методы измерения мембранного потенциала
|
из
5.00
|
Одна из важнейших функций биологической мембраны - генерация и передача биопотенциалов. Это явление лежит в основе возбудимости клеток, регуляции внутриклеточных процессов, работы нервной системы, регуляции мышечного сокращения, рецепции. В медицине на исследовании электрических полей, созданных биопотенциалами органов и тканей, основаны диагностические методы: электрокардиография, электроэнцефалография, электромиография и другие. Практикуется и лечебное воздействие на ткани и органы внешними электрическими импульсами при электростимуляции.
В процессе жизнедеятельности в клетках и тканях могут возникать разности электрических потенциалов:
1) окислительно-восстановительные потенциалы - вследствие переноса электронов от одних молекул к другим;
2) мембранные - вследствие градиента концентрации ионов и переноса ионов через мембрану.
Биопотенциалы, регистрируемые в организме, - это в основном мембранные потенциалы.
Мембранным потенциалом называется разность потенциалов между внутренней (цитоплазматической) и наружной поверхностями мембраны:
 .
.
В дальнейшем для упрощения написания формул величину 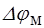 будем обозначать просто как
будем обозначать просто как 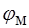 .
.
Прогресс в исследовании биопотенциалов обусловлен:
1) разработкой микроэлектродного метода внутриклеточного измерения потенциалов;
2) созданием специальных усилителей биопотенциалов (УПТ);
3) выбором удачных объектов исследования крупных клеток и среди них гигантского аксона кальмара. Диаметр аксона кальмара достигает 0,5 мм, что в 100-1000 больше, чем диаметр аксонов позвоночных животных, в том числе человека. Гигантские, в сравнении с позвоночными, размеры аксона этого проворного и ловкого головоногого моллюска имеют большое физиологическое значение - обеспечивают быструю передачу нервного импульса по нервному волокну.
Для биофизики гигантский аксон кальмара послужил великолепным модельным объектом для изучения биопотенциалов (недаром выдвигались предложения поставить памятник кальмару - животному, которому так многим обязана наука, подобно существующим памятникам лягушке в Париже и собаке под Санкт-Петербургом).
В гигантский аксон кальмара можно ввести микроэлектрод, не нанеся аксону значительных повреждений.
Стеклянный микроэлектрод (1) представляет собой стеклянную микропипетку с оттянутым очень тонким кончиком (диаметр 0,1-0,5 мкм).
Металлический электрод такой толщины пластичен, и не может проколоть клеточную мембрану, кроме того, он поляризуется. Для исключения поляризации электрода используются неполяризующиеся электроды, например, серебряная проволока (3), покрытая солью  , помещенные в раствор
, помещенные в раствор  или
или  , заполняющий микроэлектрод .
, заполняющий микроэлектрод .
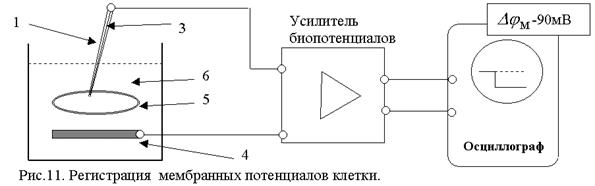 |
Второй электрод (4) для сравнения - располагается в растворе (6) у наружной поверхности клетки (5) (рис. 11). Регистрирующее устройство осциллограф.
Микроэлектродный метод дал возможность измерить биопотенциалы не только на гигантском аксоне кальмара, но и на клетках нормальных размеров: нервных волокнах других животных, клетках скелетных мышц, клетках миокарда и других.
Другим эффективным методом измерения потенциала мембраны стал метод локальной фиксации потенциала ("Patch Clamp"). Суть метода заключается в том, что микроэлектрод тонким концом, имеющим диаметр 0,5-1 мкм, присасывается к мембране таким образом, чтобы в его внутренний диаметр попал ионный канал. Тогда, используя схему фиксации потенциала, можно измерять токи, которые проходят только через одиночный канал мембраны, а не через все каналы одновременно, как это происходит при использовании стандартного метода фиксации потенциала, описанного выше.
Элемент Нернста
Природа биоэлектрогенеза была понятна лишь после создания теории электрической диссоциации (С.Аррениус 1887г.). Опыт многих исследователей убеждает, что в живых тканях, которые могут рассматриваться как растворы электролитов, генерация потенциала связана с неравномерным распределением ионов.
Самой простой и адекватной моделью является элемент Нернста (Рис.12.). Сосуд, в котором находится растворы одной и той же соли, но разной концентрации, разделен пористой перегородкой (C1 > C2). Вследствие разницы концентрации, ионы диффундируют, причем один из ионов, как правило, катион диффундирует быстрее и более разбавленный раствор приобретает знак катиона. По обе стороны перегородки образуется разность потенциалов, которую называют диффузионной разностью потенциалов.
При этом ЭДС, возникающая в концентрационном элементе, образованном раствором одной соли, определяется из уравнения Нернста.
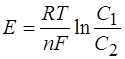
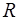 - газовая постоянная (R=8,316 Дж/к-моль),
- газовая постоянная (R=8,316 Дж/к-моль),  - абсолютная температура,
- абсолютная температура,
 - число Фарадея (96500 кл/моль), n- валентность, С1 и С2 молекулярные концентрации ионов по обе стороны мембраны.
- число Фарадея (96500 кл/моль), n- валентность, С1 и С2 молекулярные концентрации ионов по обе стороны мембраны.
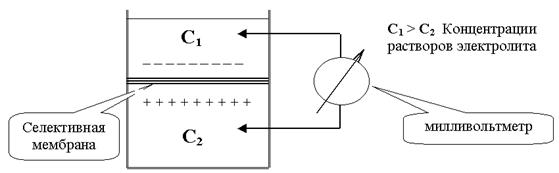
В реальных условиях, в клетке, разница скорости движения анионов и катионов обусловлена присутствием полупроницаемой мембраны. Без нее ЭДС концентрированного элемента быстро падает. Поэтому потенциалы, возникающие в таком элементе, называются не диффузионными, а мембранными.
 |
Теория электролитической диссоциации была впервые применена и объяснена в механизме биоэлектрогенеза В.Ю.Чаговцем (1896) слушателям ВМА, в дальнейшем развил учение Ю.Бернштейн и лауреаты Нобелевской премии (Ходжкин, Кац, Хакси).
Потенциал покоя.
Установлено, что цитоплазма клеток существенно отличается по химическому составу от внеклеточной жидкости, находящейся в межклеточном пространстве (рис. 13.)
| Ионы | Среда | |
| Внеклеточная | Внутриклеточная | |
| K+ | ||
| Na+ | ||
| Cl- |
Рис. 13. Относительная концентрация ионов внутри и вне клетки.
Разность ионных концентраций создает силу, стремящуюся выровнять содержание ионов, но мембрана препятствует этому процессу. Мембраны обеспечивают избирательную проницаемость для различных ионов. Вследствие этого в тканях наблюдается неодинаковая скорость диффузии через клеточные мембраны катионов и анионов, что при наличии концентрационных градиентов служит непосредственной причиной возникновения мембранных потенциалов. Их называют биопотенциалами.
Таким образом, источником электричества в живой ткани служит концентрационный элемент, образованный растворами электролита, имеющего неодинаковую концентрацию в цитоплазме и межклеточной жидкостью. При рассмотрении живых тканей в качестве концентрационного элемента необходимо учитывать диффузию через клеточную мембрану не одного иона, а всех, концентрации которых неодинаковы внутри и вне клетки. Это обстоятельство учитывает уравнение Гольдмана-Ходжкина-Катца.
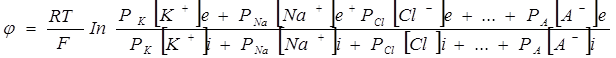 (3.3)
(3.3)
в числителе выражения, стоящего под знаком логарифма, представлены концентрации 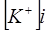 ,
, 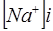 , но
, но 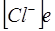 , а в знаменателе -
, а в знаменателе - 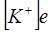 ,
,  , но
, но 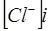 , так как ионы хлора отрицательно заряжены.
, так как ионы хлора отрицательно заряжены. 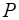 - проницаемость мембраны, зависящая от определенных условий.
- проницаемость мембраны, зависящая от определенных условий.
Уравнение Гольдмана позволяет рассчитать разность потенциалов, существующую в разных клетках между цитоплазмой и межклеточной средой в покое и при возбуждении.
Потенциал покоя - стационарная разность электрических потенциалов, регистрируемая между внутренней и наружной поверхностями мембраны в невозбужденном состоянии.
Потенциал покоя определяется разной концентрацией ионов по разные стороны мембраны и диффузией ионов через мембрану. Результаты расчетов приблизительно совпадают с величинами мембранных потенциалов, полученными опытным путем. Проанализируем уравнение Гольдмана в состояние покоя:
В состоянии покоя проницаемость мембраны для ионов 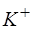 значительно больше, чем для
значительно больше, чем для 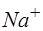 , и больше, чем для
, и больше, чем для  :
:
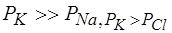
Для аксона кальмара, например,
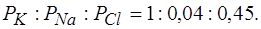 Из уравнения Гольдмана получим уравнение Нернста для мембранного потенциала покоя:
Из уравнения Гольдмана получим уравнение Нернста для мембранного потенциала покоя:
 ;
; 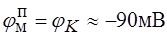
Таким образом, уравнение Нернста - частный случай уравнения Гольдмана. При концентрации в клетке 20 мМ и аксоплазме 400 мМ рассчитанное значение 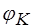 равно - 95мВ. Действительно, в опытах на аксонах регистрируют потенциалы покоя около - 90мВ. Следовательно, в состоянии покоя мембранный потенциал (ПП) равен равновесному электрохимическому потенциалу по . Важно понять, что ПП возникает за счет более быстрой диффузии по сравнению с . Разница между
равно - 95мВ. Действительно, в опытах на аксонах регистрируют потенциалы покоя около - 90мВ. Следовательно, в состоянии покоя мембранный потенциал (ПП) равен равновесному электрохимическому потенциалу по . Важно понять, что ПП возникает за счет более быстрой диффузии по сравнению с . Разница между 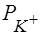 и
и 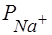 определяет направление катионного потока, создающего ПП на мембране; катионный поток (ток положительных зарядов) направлен из клетки наружу (выходящий ток) (рис.14).
определяет направление катионного потока, создающего ПП на мембране; катионный поток (ток положительных зарядов) направлен из клетки наружу (выходящий ток) (рис.14).

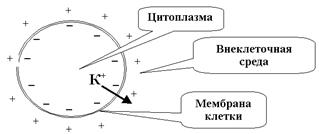 Соотношение концентраций ионов внутри и снаружи клетки определяет амплитуду ПП. Поток ионов из клетки создает разность электрических потенциалов между внеклеточной средой и цитоплазмой в условиях, когда вход в клетку и выход из нее органических анионов (что могло бы компенсировать нарушения электронейтральности от потери ) исключены свойствами покоящейся мембраны, в этой ситуации на мембране создается двойной электрический слой (снаружи - катионы, главным образом , внутри - анионы, главным образом органических кислот), препятствующий дальнейшему выходу ). Итак, все клетки в состоянии покоя характеризуются определенной степенью поляризации. Клеточная мембрана всегда заряжена. ПП в различных клетках различен и достигает нескольких десятков милливольт. У аксона кальмара ПП
Соотношение концентраций ионов внутри и снаружи клетки определяет амплитуду ПП. Поток ионов из клетки создает разность электрических потенциалов между внеклеточной средой и цитоплазмой в условиях, когда вход в клетку и выход из нее органических анионов (что могло бы компенсировать нарушения электронейтральности от потери ) исключены свойствами покоящейся мембраны, в этой ситуации на мембране создается двойной электрический слой (снаружи - катионы, главным образом , внутри - анионы, главным образом органических кислот), препятствующий дальнейшему выходу ). Итак, все клетки в состоянии покоя характеризуются определенной степенью поляризации. Клеточная мембрана всегда заряжена. ПП в различных клетках различен и достигает нескольких десятков милливольт. У аксона кальмара ПП  -85мВ, у нервных и мышечных волокон ПП -90мВ. Незначительная на первый взгляд ПП величина создает высокий градиент потенциала (напряженность) на плазматической мембране, т.к. толщина ее всего 10нм. Следовательно,
-85мВ, у нервных и мышечных волокон ПП -90мВ. Незначительная на первый взгляд ПП величина создает высокий градиент потенциала (напряженность) на плазматической мембране, т.к. толщина ее всего 10нм. Следовательно, 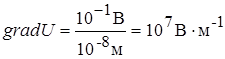 это значительная напряженность электрического поля (силовая характеристика). Особенно важно, что электрическое поле мембранного потенциала покоя поддерживает закрытое состояние так называемых активационных ворот (см. раздел 1.4.).
это значительная напряженность электрического поля (силовая характеристика). Особенно важно, что электрическое поле мембранного потенциала покоя поддерживает закрытое состояние так называемых активационных ворот (см. раздел 1.4.).
Этим и обеспечивается состояние покоя и готовность клетки к возбуждению. Избирательная проницаемость плазматической мембраны всякой клетки контролирует стабильное разобщение катионов и анионов, вследствие чего на ней стойко поддерживается высокий градиент потенциала и тем самым плазматическая мембрана генерирует ЭДС. Эта ЭДС направлена против причины, ее вызывающей, и противодействует дальнейшему разобщению зарядов на мембране, т.е. выравниванию концентраций между цитоплазмой и межклеточной средой. Существование ионных градиентов на мембранах связано также с работой активного транспорта, и является энергоемким процессом.
Нарушение биоэнергетических процессов в клетке и работы  АТФазы приводит к уменьшению
АТФазы приводит к уменьшению  .
.
Повреждение клеточной мембраны приводит к повышению проницаемости клеточных мембран для всех ионов: к повышению и  и
и  и
и 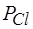 . Вследствие уменьшения различия проницаемостей, абсолютное значение мембранного потенциала
. Вследствие уменьшения различия проницаемостей, абсолютное значение мембранного потенциала 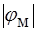 снижается.
снижается.
Для сильно поврежденных клеток еще меньше, но сохраняется отрицательный мембранный потенциал за счет содержащихся в клетке полианионов - отрицательно заряженных белков, нуклеиновых кислот и других крупных молекул, не могущих проникнуть через мембрану (доннановский потенциал). ПП отчасти экономит затраты свободной энергии на поддержание градиентов на биомембранах и препятствует их выравниванию. В этом одна из сторон физиологической роли ПП. Второй существенной стороной является то, что именно наличие ПП живой ткани способствует ее возбудимости!
Потенциал действия
Все клетки возбудимых тканей при действии различных раздражителей достаточной силы переходят в состояние возбуждения. Возбудимость - это способность клеток к быстрому ответу на раздражение, проявляющаяся через совокупность физических, физико-химических процессов и функциональных изменений. Обязательным признаком возбуждения является изменение электрического состояния клеточной мембраны. Опыт показывает, что возбужденный участок клетки становится электроотрицательным по отношению к невозбужденному, что указывает на перераспределение ионов в возбужденном участке. Оно имеет временный характер.
Общее изменение разности потенциалов между клеткой и средой, происходящее при возбуждении, называется потенциалом действия. Бернштейн полагал, что при возбуждении мембрана становится проницаемой для всех ионов. Новые градиенты исчезают, разность потенциалов на мембране уменьшается до нуля. Однако при измерении микроэлектродом оказалось, что разность потенциалов не только падает до нуля, но и продолжает изменяться дальше и достигает значительной величины.
Современная теория потенциала действия базируется на данных, полученных методами внутриклеточного отведения, фиксации напряжения на мембране и использованием радиоактивных изотопов. Потенциал действия (ПД), или потенциал возбуждения нервных клеток (волокон), возникает в ответ на достаточное по силе раздражение. ПД - очень быстрый, кратковременный электрический процесс, поэтому для его регистрации необходим катодный осциллограф с широкополосным усилителем.
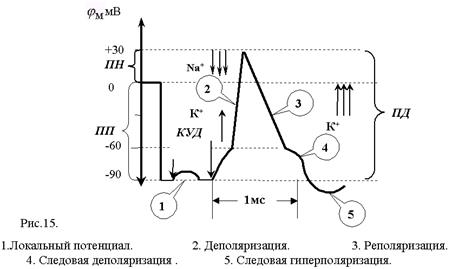
Классическое исследование параметров и механизма ПД проделано на гигантском аксоне кальмара с внутриклеточным раздражением и отведением внутриклеточного потенциала. В это нервное волокно (диаметром 0,5-1 мм) вводили на всю его длину два тончайших проволочных электрода. Один из них был раздражающим: через него в волокно подавали толчки тока того или иного направления, другой регистрировал электрический потенциал. При подаче короткого и слабого толчка выходящего тока внутриклеточный электрод регистрировал кратковременное падение МП, по форме и силе соответствующее толчку тока, но со сглаженными передним и задним фронтами, что определяется емкостью мембраны. Это так называемый локальный потенциал. Локальным он называется потому, что и в экспериментальных, и в естественных уровнях он не распространяется далеко. При усилении стимула и достижении порога раздражения, т.е. критического уровня деполяризации (КУД), возникает потенциал действия (рис.15). При толчке входящего тока любой величины образуется только электротонический
потенциал.
В потенциале действия различают пик (спайк) и следовые потенциалы.
Пик ПД представляет собой кратковременную инверсию (изменение знака на положительный) внутриклеточного потенциала. Он имеет очень быструю восходящую фазу и несколько более медленный спад. Общая длительность пика в данном объекте составляет около 3мс, амплитуда пика 120мВ, т.е. превышает МПП (90 мВ) на 30 мВ. Эту разницу называют овершутом или амплитудой потенциала инверсии.
Вслед за пиком ПД регистрируются значительно более слабые и длительные отрицательный и далее положительный следовой потенциалы (рис.15).
|
Потенциал действия имеет стандартные амплитуду и временные параметры, не зависящие от силы стимула, вызвавшего данный ПД (правило "все или ничего").
Потенциал действия - это электрический феномен, возникающий на плазматической мембране. Практически нормальный ПД возникает и в перфузируемом гигантском аксоне, лишенном аксоплазмы, при электрической стимуляции его мембраны.
Причиной развития ПД является вызываемое критической деполяризацией мембраны открытие ее натриевых и калиевых каналов. Каналы, открываемые электрическим стимулом, называют потенциалзависимыми.
Открытие потенциалзависимых каналов приводит к пассивному движению соответствующих ионов по их электрохимическим градиентам.
Вход ионов в клетку обеспечивает восходящую фазу пика ПД, т.е. деполяризацию и инверсию потенциала на мембране, а несколько запаздывающий выход ионов участвует в создании нисходящей фазы пика - реполяризации.
При развитии пика ПД отношение  становится
становится 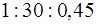 (в покое оно составляет
(в покое оно составляет 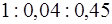 ). Связь развития пика ПД с током доказывается прямой зависимостью амплитуды ПД от электрохимического градиента на мембране и достоверным переходом меченого изотопа
). Связь развития пика ПД с током доказывается прямой зависимостью амплитуды ПД от электрохимического градиента на мембране и достоверным переходом меченого изотопа 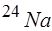 из среды в клетку при его возбуждении, причем в количестве, пропорциональном числу ПД. Связь нисходящей фазы ПД с током доказывается зависимостью хода этой фазы от электрохимического градиента на мембране.
из среды в клетку при его возбуждении, причем в количестве, пропорциональном числу ПД. Связь нисходящей фазы ПД с током доказывается зависимостью хода этой фазы от электрохимического градиента на мембране.
Подробный анализ изменений мембранной проницаемости для ионов и , основанный на измерениях токов этих ионов, стал возможным благодаря использованию методики фиксации (кламп) электрического потенциала мембраны. С помощью этих опытов было убедительно доказано, что быстрый направленный внутрь ток - это натриевый ток. Медленный задержанный ток - это калиевый ток. Быстрый входящий натриевый и медленный выходящий калиевый ток возникают практически одновременно, но ток натрия быстрее развивается, достигая своего "потолка". Если мембранный потенциал возвращается к величине, характерной для покоя, то ток натрия исчезает в 10 раз быстрее, чем ток калия, т.е. в массе натриевые каналы мембраны и активируются (деполяризацией), и деактивируются (реполяризацией) быстрее, чем калиевые. Активация каналов определяется открытием их активационных ворот (соответствующим изменением конформации неких макромолекул), а инактивация - закрытием этих же ворот. Ионные токи формируют фазы пика ПД.
 | |||
 | |||
В процессе развития ПД действуют многие факторы, связанные прямыми и обратными связями. Например, на восходящей фазе ПД действует система факторов с положительной обратной связью (рис.16).
Здесь деполяризация увеличивает 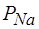 +, а последняя порождает ток
+, а последняя порождает ток 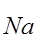 +, усиливающий деполяризацию. В силу этих отношений ток
+, усиливающий деполяризацию. В силу этих отношений ток 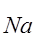 + и восходящая фаза ПД развиваются с ускорением, а амплитуда ПД быстро достигает некоторого максимума, более или менее приближенного к
+ и восходящая фаза ПД развиваются с ускорением, а амплитуда ПД быстро достигает некоторого максимума, более или менее приближенного к  +.
+.
На спаде ПД при реполяризации для тока 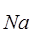 +действует та же система факторов, но с обратными знаками эффектов, а для калиевого тока - система факторов с отрицательной обратной связью.
+действует та же система факторов, но с обратными знаками эффектов, а для калиевого тока - система факторов с отрицательной обратной связью.
В итоге спад тока 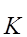 + несколько затягивается.
+ несколько затягивается.
Отрицательный следовой потенциал связан с остаточным током +, положительный следовый потенциал с остаточным током +, но главным образом с электрогенной работой натрий-калиевого насоса мембраны, активизируемого накоплением под мембраной (из-за тока Na+ ПД), а - в межклеточных щелях. В некоторых нервных клетках сразу вслед за пиком развивается довольно значительный краткий положительный следовый потенциал. Его называют андершут, он создается остаточным током K+. Расход ионов на пик одного проводящегося ПД в гигантском аксоне кальмара очень мал. Например, расход внутриаксонального при этом приблизительно равен одной миллионной доле внутреннего калиевого запаса.
Пик ПД - весьма экономичный сигнал, практически не нарушающий ионных градиентов на мембране, энергией которых он питается.
Ионные градиенты на мембране - это "пружина", энергии которой может хватить на  ПД без подзарядки. Но для длительной работы нервной клетки ионные градиенты нужно восстанавливать, что и обеспечивает работа натрий-калиевого насоса мембраны.
ПД без подзарядки. Но для длительной работы нервной клетки ионные градиенты нужно восстанавливать, что и обеспечивает работа натрий-калиевого насоса мембраны.
Рассмотрим локальный подпороговый ответ (ЛО) в нервной клетке. Этот ответ обладает в основном тем же механизмом, что и ПД. Его восходящая фаза определяется входящим током Na, а нисходящая - выходящим током .
Амплитуда ЛО пропорциональна силе раздражителя, а не стандартна, как у ПД, т.е. он не подчиняется правилу "все или ничего".
Рассчитаем равновесный электрохимический потенциал для натрия при возбуждении:
PNa+: PK+: PCl+=30:1:0,45
т.е. по сравнению с невозбужденным состоянием при возбуждении коэффициент проницаемости возрастает в 750 раз, т.к.  , уравнение Гольдмана преобразуется в уравнение Нернста, и по нему можно рассчитать равновесный потенциал по натрию. Расчет по формуле Нернста:
, уравнение Гольдмана преобразуется в уравнение Нернста, и по нему можно рассчитать равновесный потенциал по натрию. Расчет по формуле Нернста:


В опытах было доказано, что этот равновесный потенциал совпадает с амплитудным значением потенциала инверсии. ПИ всегда положителен, но меньше по абсолютной величине, чем потенциал покоя. ПД равен сумме ПП и ПИ.
Натриевый механизм потенциала действия подтверждают опыты, связанные с изменением концентрации натрия во внешней среде. При удалении ионов натрия из внеклеточной среды приводит к тому, что при стимуляции нервной клетки ПД не
возникает.
Характерные свойства потенциала действия:
1. Наличие порогового значения деполяризующего потенциала.
2. Закон "все или ничего", т.е., если деполяризующий потенциал больше порогового, развивается потенциал действия, амплитуда которого не зависит от амплитуды возбуждающего импульса и нет потенциала действия, если амплитуда деполяризующего потенциала меньше пороговой.
3. Наличие периода рефрактерности, невозбудимости мембраны во время развития потенциала действия и остаточных явлений после снятия возбуждения.
4. Резкое уменьшение сопротивления мембраны в момент возбуждения (у аксона кальмара от 0,1 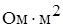 в покое до 0,0025 при возбуждении).
в покое до 0,0025 при возбуждении).
Итак, генерация потенциала действия в возбужденных мембранах возникает под влиянием различных факторов и сопровождается в первую очередь повышением проводимости клеточной мембраны для ионов натрия, входом их внутрь клетки, что приводит к деполяризации клеточной мембраны и появлению локального ответа. Этот процесс может достигнуть критического уровня деполяризации, после чего проводимость мембраны для натрия увеличивается до максимума, мембранный потенциал при этом приближается к натриевому равновесному потенциалу. Приблизительно через десять миллисекунд происходит инактивация натриевых каналов. Увеличивается активация калиевых каналов, что приводит к увеличению выходящего калиевого тока, который в свою очередь вызывает реполяризацию и в дальнейшем восстановление потенциала покоя.
Физиологическая роль ПД в потенциалзависимых клетках не ограничивается только возбудимостью клеточной мембраны и проведением нервного импульса. Возникновение ПД сопровождает целый комплекс информационных процессов, которые развиваются в ответ на стимуляцию на фоне ПД. Среди этих процессов следует назвать такие как: 1) изменение обмена веществ, 2) саморегуляция температуры и электрического импеданса, а так же других важных физиологических констант организма.
|
из
5.00
|
Обсуждение в статье: Методы измерения мембранного потенциала |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы